






研究ニュース
【共同プレスリリース】線虫が塩味の好みを逆転するしくみを解明―神経回路は餌と共に育った環境塩濃度を記憶し、その方向へ線虫をナビゲートする―
ポイント
- C. elegans(線虫)は、餌と共に飼育された環境塩濃度を好み、その方向へ進んでゆく性質を示す。
- つまり現環境より高塩濃度下で育った線虫は塩味を好み、逆に低塩濃度育成の線虫は塩味を嫌う。
- 飼育環境に依存した塩濃度記憶による塩味の好みの逆転は、塩を検知する感覚ニューロンとその下流の介在ニューロン※1の間の結合特性の反転を導く「シナプス可塑性」※2に起因することを、シミュレーション実験を用いて実証した。
- これはヒトを含む哺乳類では観測されていない、新たなタイプのシナプス可塑性である。
研究の概要
岡山大学大学院環境生命自然科学研究科博士前期課程1年 弘中 誠勝 大学院生と、室蘭工業大学大学院工学研究科 墨 智成 教授(研究当時:岡山大学異分野基礎科学研究所 准教授)は、機械学習の一種である進化的アルゴリズム※3を適用し、線虫の塩走性※4を再現する神経回路モデルを生成することで、(1)餌と共に育てられた環境塩濃度を記憶としてコード化する仕組み、(2)コード化された塩濃度記憶を読み出し、塩嗜好性(塩味の好み)を逆転させる新たなタイプのシナプス可塑性、(3)好みの塩濃度方向へ線虫をナビゲートする神経回路メカニズムを、シミュレーション実験を用いて明らかにしました。
ヒトを含むマウスなどの学習や記憶では、興奮性シナプス結合の強度を強めたり弱めたりする、いわゆる長期増強/長期抑制と呼ばれるシナプス可塑性[1]の関与が知られています。一方、塩濃度勾配下の線虫において観測される好みの塩濃度方向へ徐々に向きを変えながら進む風見鶏※5と呼ばれる塩走性では、塩濃度変化を検知する感覚ニューロンとその下流に位置する介在ニューロンの間のシナプス結合特性を、興奮性から抑制性へ反転させる新規のシナプス可塑性により、塩嗜好性を逆転している事が、シミュレーション実験を通じて明らかとなりました。
本研究による知見は、種を超えて観測される文脈依存的走性※6を制御する神経回路において、連合学習※7と記憶の読み出しがどのように実現されるのか、その根底にあるメカニズムを解明してゆくための基盤となることが期待されます。
本研究成果は令和7年2月19日に国際科学誌 elife に掲載されました。
研究の背景
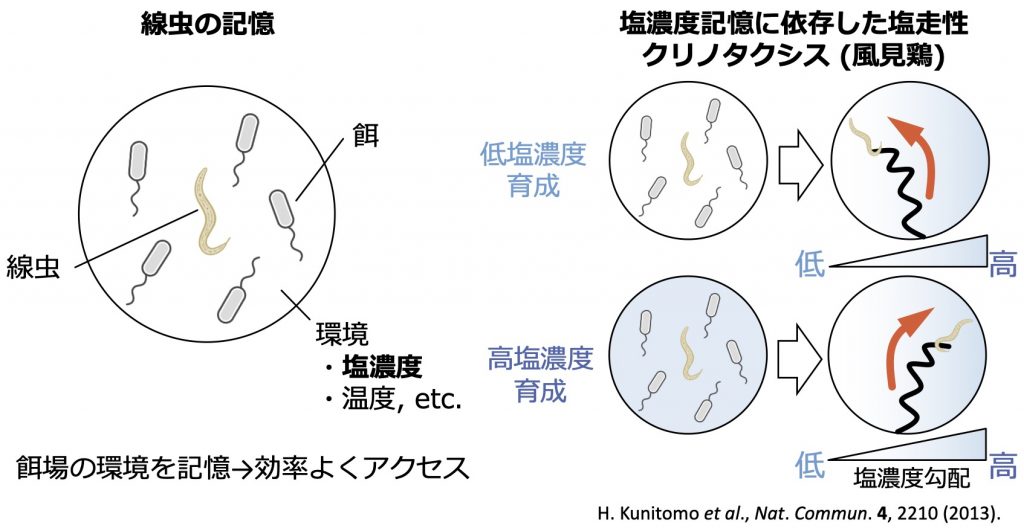
生物にとって走性は餌場に寄っていく、あるいは危険な環境を回避するために必要不可欠な生体機能であり、学習を通じてそれを適切に制御し、生存競争を勝ち抜いてゆくと考えられます。線虫 Caenorhabditis elegans は全長約1 mmの非寄生性線形動物であり、わずか302個のニューロンから成る極めてコンパクトな神経系を持ちながらも、多彩な行動を示すことから、ショウジョウバエと並び行動と神経回路の関係を理解するためのモデル生物として、精力的に研究が進められています。線虫は塩濃度勾配下におかれると、餌と共に育った環境塩濃度を好みその方向へ向かう一方、飢餓を経験したときの環境塩濃度を嫌い、逆にそれを回避することが知られています。給餌と塩濃度のいずれの条件を変えても行動が変化することから、線虫の塩走性は餌と塩濃度の関係を記憶し行動を制御する連合学習の結果として理解されます。
ところで、好みの塩濃度方向へ向かう線虫の塩走性は、大きく分けて二つの行動戦略により特徴づけられます。一つは、線虫の進行方向に沿った塩濃度勾配に応答して、急速に方向転換する「クリノキネシス」通称ピルエット、もう一つは進行方向に垂直な塩濃度勾配に応答し、塩濃度が増加(あるいは減少)する方向へ徐々に方向転換する「クリノタクシス」通称風見鶏(図1右)です。前者のクリノキネシス(ピルエット)については、給餌下での環境塩濃度に応じて好みの塩濃度(塩嗜好性)が変化し、塩濃度記憶に依存して塩走性が逆転するメカニズムが特定されています[2]。一方、後者のクリノタクシス(風見鶏)に関する塩濃度記憶の形成、並びに記憶の読み出しによる塩走性の逆転を導く神経回路メカニズムについては、実験的にも理論的にもほとんど研究が行われていませんでした。
研究の内容
線虫は解剖学研究により神経回路の地図(コネクトーム)が完全に解明されています。本研究では、クリノタクシス(風見鶏)に関与することが実験で特定されている感覚ニューロンASEL/Rおよび介在ニューロンAIZL/Rと最も強くシナプス結合を形成するニューロンを考慮した解剖学的最小神経回路[3]を出発点にし、その後明らかとなった神経電気生理学的知見[4,5]を付加条件として課した進化的アルゴリズムを適用することで、現環境より高塩濃度下で餌と共に飼育された線虫のクリノタクシス性能を最適化したベースモデル(図2左)を生成しました。その結果、ASERの下流に位置する介在ニューロンAIYL/Rとのシナプス結合は、抑制性であることが明らかとなりました。
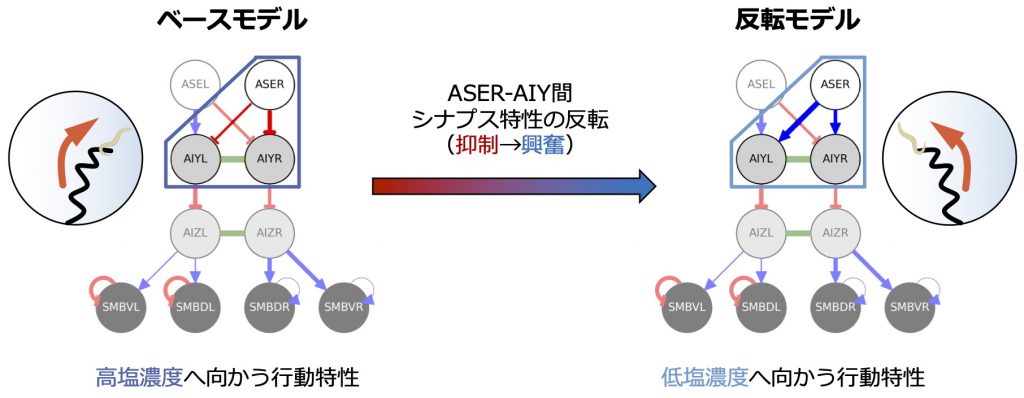
研究背景で既に述べたように、線虫の塩走性におけるもう一つの行動戦略であるクリノキネシス(ピルエット)については、給餌下での環境塩濃度に応じて塩嗜好性が変化し、塩走性の逆転を導くメカニズムが特定されています[2]。そこでは、グルタミン酸作動性感覚ニューロンASERからのグルタミン酸基礎放出量は、線虫が経験した環境塩濃度に応じて、一時的に増加/減少することが実証されています。私どもは、環境塩濃度に依存したASERによるグルタミン酸基礎放出量の変化が、クリノキネシス(ピルエット)と同様、クリノタクシス(風見鶏)においても塩濃度記憶の形成に関与するとの仮説の下、ASERの下流に位置するクリノタクシス(風見鶏)に関与する介在ニューロンAIYL/R上に発現するグルタミン酸受容体の種類を調査し、これらの知見に基づきASER-AIYL/R間シナプス特性の環境塩濃度依存性を詳細に検討しました。その結果、高塩濃度飼育の個体ではASER-AIYL/R間シナプス特性は抑制性となることが予想され、前述の付加条件下での進化的アルゴリズムにより導かれた最適化個体の結果(図2左)と一致していることが示されました。これは、本仮説の妥当性を支持する神経電気生理学的証拠であると言えます。一方、低塩濃度育成の個体では、ASERからのグルタミン酸基礎放出量の減少に伴い、ASER-AIYL/R間シナプス特性は、図2に示す様に抑制性(図2左)から興奮性(図2右)に変化する可能性が示唆されました。
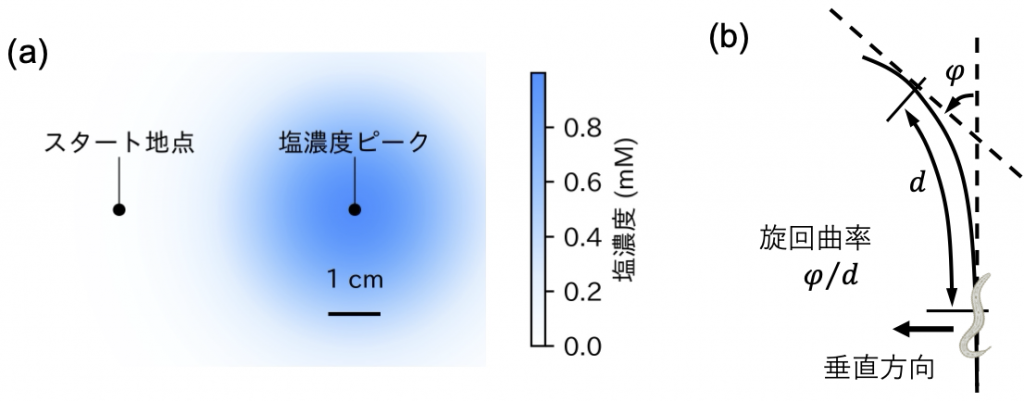
次に、本仮説に基づき導出されたASER-AIYL/R間シナプス特性の反転モデル(図2右)が、塩嗜好性の逆転を導くのか検証するために、図3aに示す様な塩濃度勾配環境下における線虫モデルのシミュレーション実験を行いました。その結果、高塩濃度育成の個体に相当するベースモデル(図2左)は、スタート地点からどの方向に出発したとしても、速やかに塩濃度のピーク方向へ旋回し、塩濃度のピーク周辺に滞在する一方(図4a)、低塩濃度育成個体に対応する反転モデル(図2右)は、様々な角度方向へ出発した個体の多くは旋回しながら塩濃度ピークと反対方向へ進んでゆくのに対し、塩濃度ピーク方向に向かって出発した一部の個体はそのままピーク方向へ進み、塩濃度ピーク周りに滞在することなくピークを通過してゆきました(図4b)。なお、図4a, 4bで示した条件を含め、様々な条件に対する両モデルによる線虫の運動軌跡は、私どもが開発した線虫シミュレーター※8(https://wormsim.streamlit.app/)を使って、実際に確認することが出来ます。
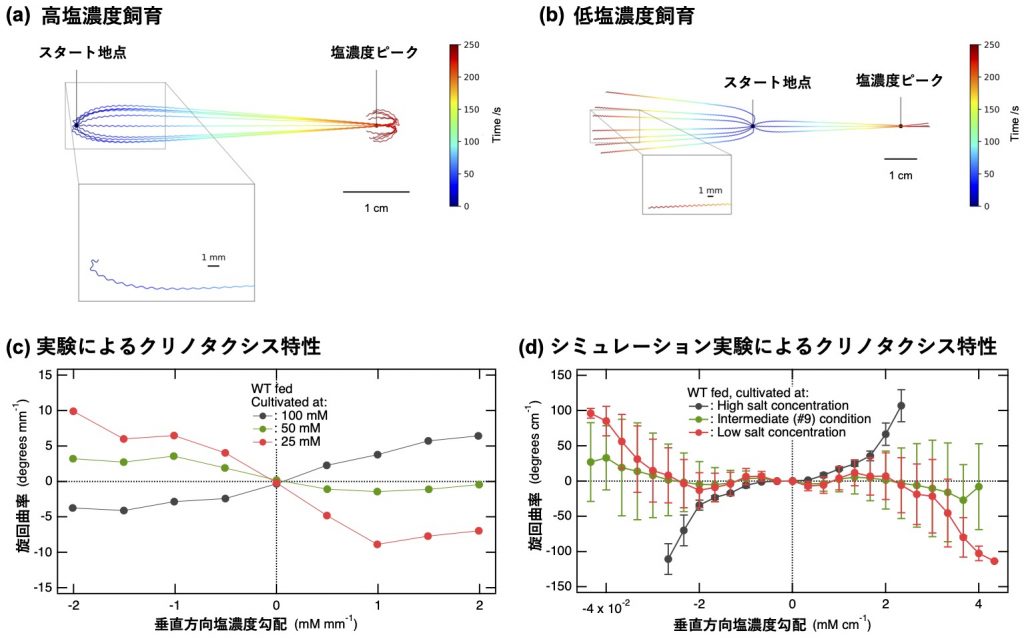
続いて両モデルによって生成される運動の軌跡が、実験で特徴付けられているクリノタクシス特性[6](図4c)と一致しているかを調べた分析結果を、図4dに示します。図4c, 4dの横軸は、線虫の進行方向に対して垂直な塩濃度勾配を示しており、縦軸は塩濃度勾配に応じて線虫がどの程度旋回したかを示す旋回曲率を示しています(図3b参照)。右肩上がりのグラフの場合、線虫は高塩濃度方向へ旋回することを示しています。実験、シミュレーション共に、黒で示した高塩濃度育成のグラフは右肩上がりになっており、一方、低塩濃度育成に対応する赤色のグラフは、左肩上がり、即ち低塩濃度方向へ旋回する事を示しています。従って、塩嗜好性の逆転が反転モデルにより上手く再現出来ていることを示しています(図4d)。緑色のグラフは現濃度と経験した環境塩濃度がほぼ等しい場合に対応しており、塩濃度勾配に対する応答が小さい事を示しています。以上から、給餌下における環境塩濃度記憶は、ASERによるグルタミン酸基礎放出量の一時的な変化として神経回路にコード化され、ASER-AIYL/Rニューロン間結合のシナプス可塑性を通じて塩濃度記憶が読み出され、飼育環境塩濃度に依存したクリノタクシスを実現していることが明らかとなりました。
ところで、飢餓状態にある線虫が採用する生存戦略として、塩濃度勾配に関係なく現在の位置から遠く離れた餌場を求めて広範囲を探索する分散行動が知られており、これがドーパミンシグナル伝達による運動ニューロンSMBの機能阻害を導くことが実験により特定されています[7]。ベースモデルおよび反転モデルにおけるSMB阻害の影響をシミュレーション実験により調査した結果、分散行動並びに実験で観測されている強く抑制されたクリノタクシス特性を再現することが示されました。これらの結果から、線虫の神経回路は、クリノタクシスに関与する最上層、すなわち環境塩濃度の影響を直接検知する感覚ニューロンASER、および最下層に位置する行動を制御する運動ニューロンSMBのみからのシグナル伝達を調整するという、考えられ得る最もシンプルな方法で、餌と塩の連合学習による行動制御を行なっている可能性が示唆されました。
今後の展開
本研究による知見は、例えばゼブラフィッシュの光走性やショウジョウバエ幼虫の走性など、種を超えて観測される文脈依存的走性を制御する神経回路における連合学習並びに記憶の読み出しの根底にあるメカニズムを解明してゆくための基盤となると期待されます。
参考文献
- [1] 岡山大学プレスリリース「なぜ私たちの脳は学習や記憶をすることができるのか~海馬興奮性シナプスにおける長期増強/長期抑制発現機構の統一的理解~」令和2年9月4日
- [2] H. Sato et al., Cell Reports 35, 109177 (2021).
- [3] E.J. Izquierdo et al., PLoS Comput. Biol. 9, e1002890 (2013).
- [4] Z. Li et al., Cell 159, 751 (2014).
- [5] A. Matsumoto et al., Proc. Natl. Acad. Sci. USA 121, e2310735121 (2024).
- [6] H. Kunitomo et al., Nature Communications 4, 2210 (2013).
- [7] A. Oranth et al., Neuron 100, 1414 (2018).
発表者
| 弘中 誠勝 | 岡山大学大学院環境生命自然科学研究科 | 博士前期課程1年 |
| 墨 智成 |
室蘭工業大学大学院工学研究科しくみ解明系領域 研究当時:岡山大学異分野基礎科学研究所 |
教授 准教授 |
論文情報
- 論文名:A neural network model that generates salt concentration memory-dependent chemotaxis in Caenorhabditis elegans
- 雑誌:elife
- 著者名:Masakatsu Hironaka, Tomonari Sumi
- DOI:10.7554/eLife.104456
研究助成
本研究成果は、令和6年度第46回両備檉(てい)園記念財団 生物学研究助成金、並びに科学研究費助成事業(23K23156)により助成を受けています。
用語解説
- ※1 介在ニューロン
感覚ニューロンや運動ニューロンと接続するニューロンで、神経回路の中継点に相当し、感覚入力による情報を処理/統合し、運動ニューロンへ伝達する役割を担います。 - ※2 シナプス可塑性
神経回路を形成するニューロン同士をつなぐシナプスが、経験や学習によって変化する現象で、学習や記憶、運動性など、神経回路の機能に関わる重要な性質です。ヒトを含む哺乳類の脳におけるシナプス可塑性では、シナプス結合の強度を増強あるいは抑制することで学習や記憶の形成が行われますが、結合特性を興奮から抑制へ反転する(あるいはその逆の)シナプス可塑性は、ヒトを含む哺乳類では観測されていません。一方、線虫ではシナプス特性の反転を導く新たなタイプのシナプス可塑性が発見されています。 - ※3 進化的アルゴリズム
機械学習の一種で、生物の進化を模倣して最適解を探索するアルゴリズムです。自然淘汰と進化のプロセスに着想を得ており、何世代にもわたって候補解の母集団を繰り返し改善することで、複雑な最適化問題を解くことが出来ます。ここでは、線虫が高塩濃度へ向かう行動性能を向上させることで、最適化された神経回路モデルを生成しています。 - ※4 塩走性
生物が化学物質の濃度差を検知して一定の方向に移動する現象「化学走性」の一種。線虫は餌となる微生物が発する塩の濃度変化を検知して、餌を求めてその塩濃度へ向かう「塩走性」を示す。 - ※5 風見鶏
線虫の塩走性は、線虫の進行方向の塩濃度勾配に反応して進行方向を反転するklinokinesis(クリノキネシス)、別名ピルエットと、進行方向と垂直な塩濃度勾配に反応して、進行方向を徐々に変化させるklinotaxis(クリノタクシス)、別名風見鶏によって構成されている。風見鶏では塩味を好む場合、進行方向と垂直な方向の塩濃度が増加する方向へ徐々に進行方向を変化させ、高塩濃度方向へ向かって旋回してゆく(図1右参照)。 - ※6 文脈依存的走性
文脈依存性とは、物事を覚えるときや思い出すときの環境が一致しているほど、その物事を想起しやすいという現象で、ここでは、経験した環境に応じて行動選択が影響を受け、結果として走性が変化することを意味している。 - ※7 連合学習
動物が複数の刺激を関連付けて学習し記憶する事。例えば、ショウジョウバエに匂い刺激と同時に電気ショックあるいはショ糖をそれぞれ、嫌悪/報酬刺激として与える嗅覚連合学習では、電気ショックとペアになった匂いを忌避(きひ)(回避)し、逆にショ糖とペアの匂いを欲求するようになる。 - ※8 線虫シミュレーター
高塩濃度育成個体に対応するベースモデル(図2左)と低塩濃度育成個体に対応する反転モデル(図2右)による様々な条件下でのシミュレーション実験を行うためのWebアプリ。 URL(https://wormsim.streamlit.app/)から使用することができる。

研究に関する問い合わせ
室蘭工業大学大学院工学研究科しくみ解明系領域 教授
墨 智成(すみ ともなり)
Tel:0143-46-5724 E-mail:sumi@muroran-it.ac.jp
報道に関する問い合わせ
国立大学法人室蘭工業大学総務広報課秘書広報係
Tel:0143-46-5008
E-mail:koho@muroran-it.ac.jp
岡山大学総務・企画部広報課
Tel:086-251-7292
E-mail:www-adm@adm.okayama-u.ac.jp